
orecchio
Anatomia e medicina
Organo pari dei Vertebrati, situato ai 2 lati del capo, che ha la caratteristica funzione dell’udito e partecipa, insieme ai centri encefalici e midollari, alla regolazione dell’equilibrio statico e dinamico.
Anatomia comparata
Negli invertebrati la capacità di orientarsi nello spazio è dovuta a organi di senso statico, o statocisti, che contengono una o più concrezioni calcaree, le statoliti o otoliti. Organi di senso acustici, tuttavia non paragonabili all’o. dei Vertebrati, sono gli organi cordotonali e timpanici degli Insetti.
In tutti i Vertebrati è presente l’organo di senso statico che, negli Ittiopsidi, Anfibi Urodeli compresi, è rappresentato dall’o. interno; nei Tetrapodi, invece, la funzione stato-recettrice dell’o. si associa a quella auditiva: dagli Anfibi Anuri in poi compare l’o. medio per la trasmissione delle vibrazioni sonore dall’aria all’o. interno. L’o., nei Vertebrati terrestri, adempie perciò la funzione di organo stato-acustico. Nei Mammiferi l’o. si complica ulteriormente per la presenza dell’o. esterno, che serve a raccogliere i suoni e a farne individuare la provenienza.
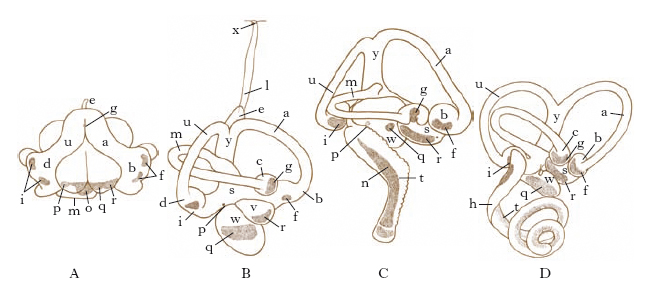
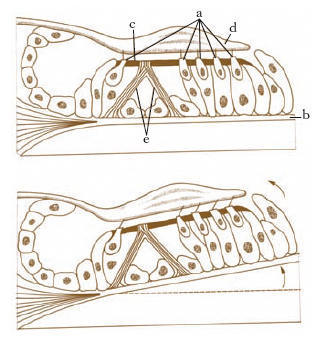
L’o. interno si sviluppa nei Vertebrati da un ispessimento ectodermico situato ai due lati del capo che, invaginandosi, si trasforma in una otocisti, o vescicola otica, che rimane in comunicazione con l’esterno attraverso il dotto endolinfatico, il quale resta pervio solo nei Selaci. Successivamente l’otocisti, in cui frattanto si è differenziata medialmente un’area di epitelio sensoriale (macula communis), subisce una costrizione in due parti: il vestibolo, o otricolo dorsale, e il sacculo ventrale, connessi da un canale sacculo-utricolare. L’area sensoriale mediale della vescicola si suddivide a sua volta in 2, dando la macula dell’otricolo e del sacculo. Quindi dalle pareti dell’otricolo si producono tre evaginazioni appiattite, gli abbozzi dei canali semicircolari, che poi si identificano come tubi semicircolari aprentisi ai due estremi nell’otricolo; quello laterale è situato nel piano orizzontale, l’anteriore e il posteriore in piani verticali. A un’estremità di ciascuno dei canali la parete si dilata in ampolle al cui interno si differenzia un’area sensoriale, la crista acustica. Nei Missiformi vi è un unico canale semicircolare e un dotto endolinfatico rudimentale; nei Petromizontiformi si hanno invece i due canali semicircolari verticali; nei Selaci sono presenti tre canali semicircolari. Mentre nei più bassi Ittiopsidi il sacculo si differenzia scarsamente, negli altri compare un’evaginazione posteriore, la lagena, con l’epitelio sensoriale. Nei Tetrapodi la lagena si sviluppa maggiormente e insieme con la papilla basilare, altra estroflessione basale del sacculo, che appare già negli Anfibi, costituisce la coclea, che è la struttura più propriamente acustica dell’o. interno dei Vertebrati. L’insieme delle varie parti e strutture forma il labirinto membranoso, ripieno di un fluido, l’endolinfa, nel quale sono sospese particelle calcaree, rappresentate da microscopici cristalli di carbonato di calcio (otoconie) o, come nei Teleostei, da uno o più grossi otoliti. Con la formazione del neurocranio cartilagineo, il labirinto membranoso è incluso in una capsula protettiva cartilaginea, la capsula otica: tra questa e quello si interpone lo spazio perilinfatico con la perilinfa. Con l’ossificazione del neurocranio, il labirinto scheletrico risulta formato dalle varie ossa otiche. Nei Rettili la coclea si allunga, nei Coccodrilli manifesta una tendenza a ripiegarsi, come negli Uccelli, finché nei Mammiferi si avvolge a spira in più piani, costituendo la rampa, o scala media, della coclea. Lo spazio perilinfatico si suddivide in 2 canali spiralati: il superiore, la scala vestibuli, e l’inferiore, la scala tympani. Nel labirinto membranoso le aree sensoriali dell’epitelio sono rappresentate dalle creste acustiche nelle ampolle dei canali semicircolari, dalle macule acustiche nell’otriculo e nel sacculo e dalle papille acustiche basilare e lagenare: quest’ultima è assente nei Mammiferi, dove la basilare costituisce l’organo del Corti della percezione dei suoni (fig. 1).
L’o. medio, presente in tutti i Tetrapodi (eccettuati i Serpenti e qualche Anfibio Apode), si origina in relazione con la prima fessura branchiale e consta della cassa del timpano, o cavità timpanica, della membrana del timpano che lo separa dall’esterno, e della tromba d’Eustachio che lo fa comunicare con la faringe. La cavità timpanica, omologa allo spiracolo degli Elasmobranchi, è situata anteriormente e al disotto della capsula periotica, che presenta in essa due aperture: la finestra rotonda (fenestra tympani), inferiore, e la finestra ovale (fenestra vestibuli), superiore, chiuse da una membrana che separa la cavità timpanica dagli spazi perilinfatici dell’o. interno. Il timpano è superficiale negli Anfibi, in molti Rettili e in alcuni Uccelli. Nella cavità timpanica, fra la finestra ovale e il timpano, è interposta una catena di ossicini formata da staffa e columella in Anfibi, Rettili e Uccelli, staffa, incudine e martello nei Mammiferi; la staffa è connessa alla membrana della finestra ovale, la columella, o il martello, alla membrana timpanica.
L’o. esterno compare come breve condotto auditivo in alcuni Rettili e Uccelli. Nei Mammiferi, dietro il meato uditivo esterno si sviluppa, eccetto che nei Cetacei, nei Sirenidi e in alcuni Pinnipedi, il padiglione auricolare o conca che raccoglie le onde sonore e le convoglia al timpano.
Anatomia umana

Nell’o. dell’uomo si distinguono o. esterno, o. medio e o. interno. Le prime due parti raccolgono le onde sonore e le trasmettono all’o. interno, nel quale sono localizzati gli organi sensitivi, sia acustici sia dell’equilibrio (fig. 2).
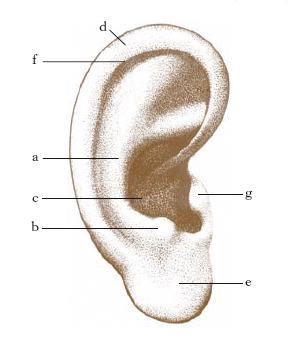
O. esterno
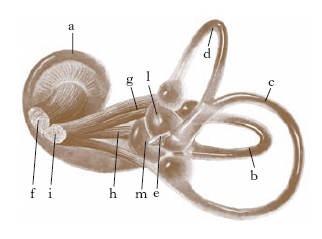
Il padiglione dell’o. ha forma concava, irregolarmente ovale, con il maggior asse verticale, e presenta una serie di rilievi e depressioni: l’elice, l’antelice, il solco dell’elice, il trago, l’antitrago e il tubercolo di Darwin (fig. 3). Nella sua parte inferiore si nota una ripiegatura cutanea detta lobo (o lobulo). Il padiglione dell’o., a eccezione del lobulo, che è costituito unicamente da tessuto molle, è formato da un sottile scheletro cartilagineo, ricoperto da un esile strato di tessuto sottocutaneo e da cute sottile. Nello spessore del sottocutaneo si trovano alcuni muscoli, che nell’uomo sono rudimentali.
Il padiglione si continua verso l’o. medio con il meato uditivo esterno, che è un condotto grossolanamente cilindrico, lungo circa 24-25 mm, che termina a fondo cieco con la membrana del timpano. La cute del meato uditivo esterno contiene le ghiandole ceruminose, affini alle ghiandole sudoripare. Nella porzione esterna il condotto uditivo poggia su uno scheletro cartilagineo, nella parte interna decorre in un canale scavato nell’osso temporale.
O. medio
L’o. medio è detto anche timpano, perché è costituito da una cavità piena d’aria a forma di tamburo situata nello spessore dell’osso temporale. La parete esterna è formata per la sua maggiore estensione da una sottile membrana fibrosa (membrana timpanica), di forma circolare, con diametro di 9-10 mm, che si inserisce su un anello osseo: la parte superiore dell’o. medio, quella che corrisponde alla parte superiore di questo anello osseo, prende il nome di loggetta degli ossicini.
La parete mediale è in rapporto con l’o. interno e vi si osservano diverse formazioni: una sporgenza ossea detta promontorio, che corrisponde al primo giro della spirale della chiocciola; due orifizi, uno di forma ovale (finestra ovale), occupato dalla base della staffa e da una membrana periostea, che corrisponde al vestibolo dell’o. interno, e uno di forma rotonda (finestra rotonda) chiuso da una lamina periostea (timpano secondario), che corrisponde all’estremità inferiore della chiocciola.
Nella circonferenza, la parte superiore è costituita da una sottile lamina ossea che separa l’o. medio dalla cavità cranica; la parte inferiore, anch’essa a volte molto sottile, lo divide dalla vena giugulare; nella parte posteriore si apre un ampio canale (canale timpanico-mastoideo) che comunica con le cellule mastoidee, mentre nella parte anteriore sbocca la tromba di Eustachio che mette in comunicazione l’o. medio con la faringe.
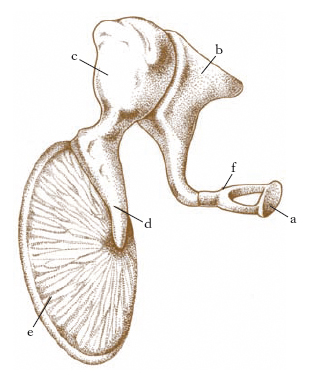
Tra la parete membranosa e la parete ossea sono disposti 3 elementi ossei, il martello, l’incudine e la staffa, che, essendo articolati tra loro, costituiscono la cosiddetta catena degli ossicini (fig. 4); di essi, il martello si inserisce sulla membrana timpanica, la staffa nella finestra ovale. Due esili muscoli (muscolo del martello, muscolo della staffa) formano l’apparato motore della catena.
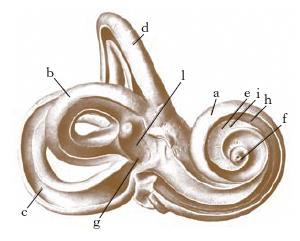

O. interno
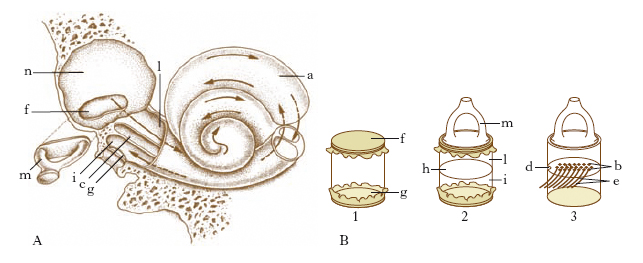
È contenuto in un sistema di cavità (coclea, vestibolo, canali semicircolari) scavate nella rocca petrosa del temporale, che nel loro insieme costituiscono il labirinto osseo (fig. 5). La parte funzionale è rappresentata dal labirinto membranoso, che a sua volta è costituito da una serie di vescicole la cui denominazione ripete quella degli elementi del labirinto osseo (fig. 6). Con criteri topografici e funzionali il labirinto viene distinto in anteriore (coclea) e posteriore (vestibolo, canali semicircolari). Tra il labirinto osseo e quello membranoso è contenuto un liquido, detto perilinfa (fig. 7); all’interno del labirinto membranoso esite un liquido analogo, detto endolinfa.
La coclea ossea è costituita essenzialmente da un condotto cilindrico, ravvolto a tre giri di spira incompleti intorno a un nucleo osseo compatto (columella o modiolo). Il modiolo alla base presenta una serie di minuscoli forellini che danno passaggio alle fibre del nervo cocleare: sono ordinati in doppia fila e nel loro insieme costituiscono il cribro spiroide della base della coclea; per tutta la sua lunghezza è percorso da una lamina, che è detta lamina spirale, perché segue le volute della coclea: congiunta come è a una membrana (membrana basilare), che la prolunga inserendosi sulla parete della coclea, costituisce un sepimento che divide il lume del condotto in due concamerazioni o rampe, dette, per i loro rapporti, rispettivamente timpanica e vestibolare. Una terza concamerazione, denominata dotto cocleare, è ricavata nell’ambito della rampa vestibolare da un altro sepimento, la membrana vestibolare di Reissner, che si inserisce da una parte sulla lamina spirale in prossimità della membrana basilare e dall’altra sulla parte esterna del tubo cocleare.
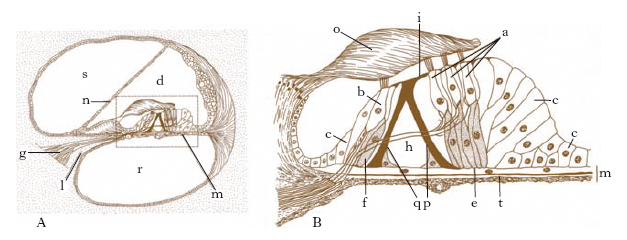
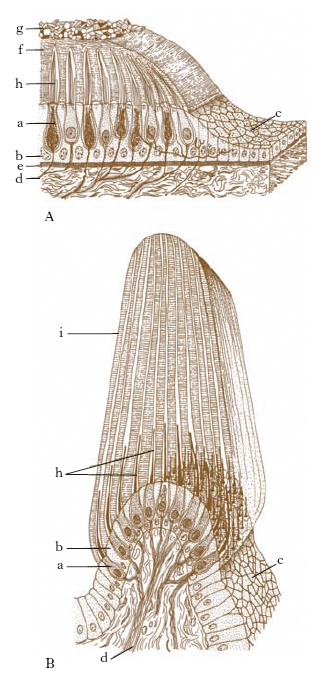
La rampa vestibolare si estende dal vestibolo all’apice della coclea, al cui livello comunica con la rampa timpanica per mezzo di un piccolo orifizio detto elicotrema; il dotto cocleare è chiuso alle estremità, ma comunica col sacculo mediante un esile canale, il dotto di Hensen. Le 3 concamerazioni sono caratterizzate dalla presenza nel loro interno di un liquido: la perilinfa nelle due rampe, l’endolinfa nel dotto cocleare (fig. 8A). In quest’ultimo, impiantato sulla membrana basilare, è allogato l’organo spirale del Corti, l’apparato specifico di ricezione delle stimolazioni uditive (fig. 8B). Alla costituzione dell’organo spirale del Corti prendono parte le fibre del nervo cocleare e un epitelio formato da cellule sensoriali e da cellule di sostegno.
Il vestibolo osseo è una cavità ovoide interposta tra la cassa del timpano, la coclea e i canali semicircolari; nella sua parete mediale i minuscoli forellini della zona cribrosa (macule cribrose) permettono alle fibre di origine del nervo vestibolare di raggiungere il meato acustico interno per unirsi al nervo cocleare, dando origine al nervo acustico; nella stessa parete si apre un particolare condotto (acquedotto del vestibolo) che accoglie il dotto endolinfatico. Il vestibolo membranoso comprende 2 distinte vescicole: l’otricolo in comunicazione coi canali semicircolari, il sacculo in comunicazione con la coclea; dalle due vescicole si dipartono due canalini, uno per ciascuna formazione, che confluendo fra loro danno origine al dotto endolinfatico, che termina a fondo cieco in contatto con la dura madre endocranica. L’otricolo e il sacculo nella loro superficie interna presentano ciascuno una piccola sporgenza biancastra, indicata col nome di macula acustica (fig. 9A), che corrisponde al punto di origine del nervo specifico, rispettivamente nervo otricolare e nervo sacculare. Sulle macule poggia il cosiddetto apparato otolitico, costituito da uno strato gelatinoso (membrana otolitica) che ingloba particolari concrezioni di calcio e di magnesio, dette otoliti.
I canali semicircolari, in numero di tre, sono distinti in superiore, posteriore e laterale e sono orientati secondo le 3 direzioni dello spazio, essendo tra loro perpendicolari; con i loro estremi sboccano nel vestibolo; il loro epitelio presenta un ispessimento (cresta acustica) analogo per significato alle macule (fig. 9B).
Fisiologia
L’o., oltre a essere l’organo dell’udito, partecipa, assieme ai centri encefalici e midollari, alla regolazione dell’equilibrio statico e cinetico. I recettori per l’udito sono localizzati nella coclea, quelli per l’equilibrio nell’otricolo, nel sacculo e nei canali semicircolari.
Funzione acustica
L’o. umano percepisce le vibrazioni sonore la cui frequenza è compresa tra 16 e 20.000 Hz. Le vibrazioni sonore sono convogliate dal padiglione auricolare nel condotto uditivo esterno e fanno vibrare la membrana timpanica, che fa entrare in azione il sistema di leve rappresentato dalla catena degli ossicini; quest’ultima non si limita a trasmetterle con la staffa, suo segmento terminale, e attraverso la finestra ovale alla perilinfa, ma, per le caratteristiche geometriche, le modifica, riducendone l’ampiezza e aumentandone l’intensità, adattando così l’impedenza acustica specifica dell’aria, da cui provengono le onde sonore, a quella del liquido della coclea, dove le onde si trasmettono. Altre modalità di trasmissione delle onde sonore all’o. interno sono quella per via ossea e quella rappresentata dalle variazioni di pressione dell’aria contenuta nell’o. medio, prodotte dalle vibrazioni del timpano e trasmesse alla chiocciola dalla membrana (timpano secondario) che occlude la finestra rotonda. Le onde di pressione indotte nella perilinfa dalle vibrazioni della staffa, causa la inestensibilità delle pareti ossee, si propagano lungo la rampa vestibolare in direzione dell’apice, per ridiscendere, attraverso l’elicotrema, nella rampa timpanica e investono anche le pareti membranose del dotto cocleare trasmettendosi all’endolinfa (fig. 10): se la loro frequenza è bassa, l’onda di pressione si propaga fino in vicinanza dell’apice; se elevata, raggiunge la prossimità della base della chiocciola. In seguito all’attivazione della catena ossiculare si verificano variazioni di pressione nei liquidi labirintici (perilinfa ed endolinfa) indotte dai movimenti della staffa. Le onde che si vengono a determinare deformano in modo proporzionale allo stimolo la partizione cocleare (organo del Corti: membrana tectoria, membrana basilare ecc.; fig. 11). Si crea così un’onda viaggiante che si propaga verso l’elicotrema. La deformazione indotta nella partizione cocleare è uno stimolo adeguato ad attivare anche le strutture dell’organo del Corti meno sensibili alle variazioni pressorie: le cellule ciliate. È qui che mediante variazioni di potenziali di membrana si crea un flusso elettrico capace di liberare i mediatori sinaptici verso le terminazioni afferenti del 1° neurone uditivo. La successiva fase della percezione uditiva è di codificazione dell’impulso, diversa a seconda della frequenza del segnale in entrata. A tal fine il 1° neurone ha a disposizione fondamentalmente 3 meccanismi: fibre nervose che presentano una propria specificità di frequenza; possibilità di variare la cadenza dell’impulso neurale; attivazione in parallelo di più fibre nervose.
Gli studi di elettrofisiologia uditiva applicati poi alla clinica (audiometria a risposte elettriche) permettono di visualizzare e quantificare i fenomeni di trasduzione meccanico-elettrica (potenziale microfonico cocleare e potenziale di sommazione) e di attivazione delle fibre del primo neurone (potenziale di azione del nervo uditivo). Tali mezzi di indagine, che si aggiungono alle prove di audiometria tonale e vocale e di impedenziometria, offrono al medico la possibilità di identificare in caso di sordità neurosensoriale la sede precisa della lesione.
Funzione labirintica
Gli stimoli propri dell’apparato labirintico sono di natura meccanica. Per i canali semicircolari sono dati dagli spostamenti dell’endolinfa rispetto alla parete dei canali semicircolari, tardando questa, per la sua inerzia, a seguire i movimenti più rapidi dei canali. Tale meccanismo spiega come gli impulsi che da questo livello arrivano alla corteccia si riferiscono all’entità dei movimenti del capo e alla relativa accelerazione angolare (funzione cinetica) e non alla posizione statica del capo. Gli stimoli appropriati del sacculo e dell’otricolo sono costituiti invece dalle sollecitazioni meccaniche che l’apparato otolitico esercita sulle cellule epiteliali delle macule, o col loro peso o con la trazione, quando i movimenti del capo tendono a modificare i rapporti tra otoliti e macule. Questa loro funzione è detta statocinetica, perché informa la corteccia non solo sulla posizione del capo rispetto alla forza di gravità, ma anche sulle variazioni di intensità e di direzione di qualsiasi altra forza che imprima un’accelerazione e produca un moto, lineare o rotatorio. Le connessioni che il nucleo del nervo vestibolare ha con altri centri dell’asse encefalico (centri mesencefalici, protuberanziali, bulbari), del cervelletto e del midollo spinale, permettono all’apparato labirintico di partecipare alla regolazione del tono e dell’attività posturale dei muscoli del tronco e degli occhi e di intervenire sulla funzione cerebellare (➔ equilibrio).
Patologia
L’o., nelle sue varie parti, può essere sede di numerose anomalie e affezioni. A carico del padiglione e del condotto uditivo esterno si possono osservare malformazioni (microtia, atresia del condotto ecc.), lesioni traumatiche e affezioni cutanee comuni agli altri distretti corporei (come eczemi, micosi, foruncoli). Disturbi di tipo particolare si possono avere in caso di occlusione del condotto uditivo (per tappi di cerume, penetrazione di corpi estranei, esostosi, tumori). L’o. medio va incontro, con notevole frequenza, a processi infiammatori (➔ otite), catarrali o purulenti. Le affezioni dell’o. interno, o labirintopatie, sono causa di quadri morbosi (per es., la sindrome di Ménière) nei quali dominano i disturbi dell’udito e dell’equilibrio.
Religione
L’attributo di un’accentuata disponibilità di o. è proprio di numerose figure divine, semidivine o semplicemente mitologiche a tutti i livelli storico-religiosi come espressione diretta dell’onniaudienza, quasi sempre associata all’onniveggenza (che si manifesta nella polioftalmia) nel quadro generale dell’onniscienza. Per l’o. come segno dell’onniaudienza è generalmente usata la semplice pluralità: nell’Egitto antico a una divinità solare non nominata sono attribuiti 77 o. (insieme a 77 occhi) che per Ra raggiungono il numero di 777; nel politeismo babilonese Marduk, possessore di una sapienza di tipo magico e oracolare, si presenta anche come depositario di un’onniscienza automaticamente visiva e auditiva (nell’Enūma elīsh I, 95 Marduk ha 4 occhi e 4 o.); nel Mazdeismo sono attribuiti a Mithra 1000 o. ecc. Questa pluralità può essere sostituita da dimensioni eccezionali o da una particolare posizione degli stessi o. (Numi-Tarem dei Voguli ha o. «grandi come le foglie del nenufar»; nel pensiero eclettico taoista-confuciano a Tian, localizzato nelle altezze supreme, sono attribuiti «o. bassi»; Leza nella religione di popolazioni bantu centrali «ha o. lunghi» ecc.). La pluralità di o. è implicita nella policefalia che caratterizza alcune divinità, sia nel mondo classico (Borea, Giano ecc.) sia nell’Europa barbarica (Traci, Celti, Slavi occidentali). Sono invece escluse da ogni considerazione in proposito figure la cui policefalia si manifesta quale indicazione di mostruosità (per es., l’Idra di Lerna nella mitologia classica).
Tecnica
In elettroacustica e in acustica fisiologica, o. artificiale, ricevitore, per es. un microfono, la cui risposta acustica è simile a quella dell’o. umano e che può pertanto sostituirsi a questo nelle prove di audiometri, trasduttori elettroacustici ecc.; in particolare, o. normale, o o. assoluto, è un o. artificiale la cui risposta acustica corrisponde perfettamente all’audiogramma normale, e come tale è atto quindi a simulare il comportamento di un o. umano ‘normale’.