
occhio
Anatomia e medicina
Organo di senso per la ricezione degli stimoli luminosi, che vengono trasmessi ai centri nervosi dando origine alle sensazioni visive.
Anatomia comparata
Invertebrati
Molti Invertebrati reagiscono agli stimoli luminosi grazie alla presenza di cellule sensoriali isolate sparse sul corpo. In altri, invece, le cellule si raggruppano e la ricezione è migliorata per la comparsa di uno schermo di pigmento; questo, limitando la ricezione alla luce proveniente da una sola direzione, permette l’orientamento. Un ulteriore aumento di complessità è rappresentato dall’o. a fossetta di molti Molluschi, con le cellule sensibili disposte in un’invaginazione epiteliale; se la fossetta si chiude l’o. funziona come una camera oscura (Nautilus). La comparsa di apparati diottrici e catottrici permette, infine, una formazione delle immagini ottimale ed evita la dispersione dei raggi luminosi (Cefalopodi).
Gli Artropodi possiedono o. semplici e o. composti. Gli o. semplici (o ocelli) constano di una lente e di uno strato cellulare sensibile (retina). Negli Insetti sono tipicamente in numero di tre, situati sulla regione dorsale del capo, fra gli occhi composti. Lo stesso nome si usa anche per le macchie oculari, organi fotorecettori costituiti da poche cellule sensorie, propri di molti Invertebrati (molluschi, cnidari, nematodi, asteroidei ecc.), che, a differenza dell’o. dei Vertebrati e dei Cefalopodi e dell’o. composto degli Artropodi, non consentono la formazione di un’immagine.
Gli o. composti sono costituiti da un numero vario di elementi detti ommatidi (talvolta anch’essi chiamati impropriamente ocelli), unità sensoriali provviste di un apparato diottrico rappresentato da una cornea o corneola e da un cono cristallino. La parte sensibile, retinula, è formata da cellule sensoriali raggruppate intorno a un rabdoma. Ogni ommatidio è avvolto da un manicotto di cellule pigmentate a funzione catottrica, e a volte è presente un tapetum che riflette i raggi luminosi dal fondo dell’o., permettendo così di utilizzare anche minime quantità di luce. Il numero di ommatidi varia molto nei diversi gruppi. La parte esterna dell’o. composto è una calotta sfaccettata per la presenza di corneole, una per ogni ommatidio. L’o. composto dà una visione ‘a mosaico’; l’immagine sarà poi integrata nei centri nervosi.
Vertebrati
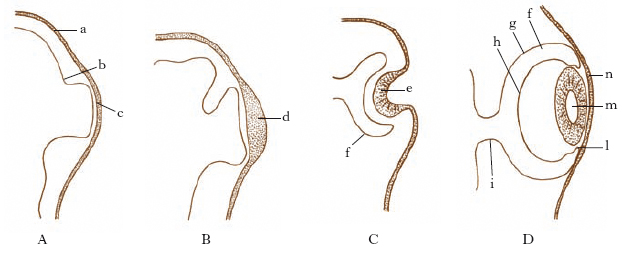
Gli o. pari dei Vertebrati si originano da due intumescenze del prosencefalo, le vescicole ottiche primarie, che successivamente si trasformano in calici o coppe ottiche, unite all’encefalo attraverso i peduncoli ottici (fig. 1). I calici sono a duplice parete: interna, che darà la retina, ed esterna, che si differenzia nell’epitelio pigmentoso retinico. Ventralmente, il calice ottico presenta la fessura coroidea che prosegue in una doccia del peduncolo dove passeranno le fibre del nervo ottico provenienti dalla retina. L’ectoderma della parete distale del calice ottico forma intanto il placode lentogeno che, successivamente, si distacca, formando la vescicola lentogena, abbozzo del cristallino. Il foglietto retinico si differenzia intanto nei tre strati di neuroni tipici della retina definitiva. Gli altri costituenti dell’o. dell’adulto sono: la sclerotica, tunica a funzione protettiva; la cornea, calotta trasparente posta nella parte anteriore del bulbo oculare; l’umore acqueo, che riempie la camera anteriore e posteriore; l’umore vitreo, semisolido, posto fra la retina e la lente, che con la cornea, il cristallino e l’umore acqueo forma l’apparecchio rifrangente (fig. 2); la coroidea, il corpo ciliare e l’iride che costituiscono le membrane uveali da cui derivano gli organi pigmentosi dell’o. dei Vertebrati.
In tutti i Vertebrati gli o. sono costruiti sul modello di una camera oscura con apparato diottrico e uno strato sensoriale, la retina. Occhi rudimentali sono presenti in organismi (Proteus, Typhlomolge, Talpa ecc.) endogei o cavernicoli. Gli o. degli Agnati viventi sono rudimentali; i Condroitti e gli Osteitti presentano una cornea molto appiattita e un cristallino sferico; nei Teleostei esiste anche il processo falciforme, caratteristica struttura muscolare che si estende dalla retina al cristallino, con funzioni di messa a fuoco. Particolari condizioni sono presentate dai pesci della famiglia Pleuronettidi, con i due o. disposti su un solo lato, destro o sinistro, del corpo, e in alcuni pesci abissali che hanno spesso grandi o. telescopici e peduncolati.
L’o. dei Sauropsidi possiede particolari strutture: il cono nei Rettili e il pettine negli Uccelli, che originano dal fondo dell’o. e sporgono nel vitreo; il cono potrebbe avere funzioni nutritive, il pettine invece potrebbe servire anche per una migliore individuazione di piccoli oggetti mobili e per l’orientamento solare (il pettine proietta la sua ombra sulla retina agendo come lo gnomone di una meridiana). La particolare acutezza visiva degli Uccelli, che normalmente possiedono grandi bulbi oculari, dipende dall’alta densità dei fotorecettori retinici (un milione di coni per mm2 nella fovea di un falco, il triplo rispetto all’uomo) e dalla velocità di accomodamento della visione. In diversi Rettili e Uccelli, inoltre, parti della sclerotica si ossificano a formare un anello di ossa sclerotiche con funzioni di irrobustimento della stessa sclera. Gli o. dei Mammiferi sono sferici e piuttosto grandi, soprattutto nelle specie acquatiche.
L’iride è un diaframma circolare con un’apertura centrale, la pupilla, generalmente circolare, ma a volte ellittica o a fenditura orizzontale o verticale. Lo strato posteriore dell’iride è ricco di pigmenti di diversi colori e il resto dell’iride è muscolare, con fascetti disposti circolarmente e radialmente intorno alla pupilla, di cui, contraendosi, fanno variare il diametro. Nei pesci Teleostei l’accomodazione si realizza con contrazioni del muscolo retrattore del cristallino, sferico e indeformabile, che può essere spostato in avanti e indietro. Nei Rettili e negli Uccelli si modifica invece la superficie anteriore del cristallino. Nei Mammiferi l’accomodazione avviene come nell’Uomo.
L’o. è variamente mobile nelle orbite grazie a muscoli oculomotori striati. Accessorie dell’o. sono le palpebre, pieghe del tegumento che proteggono il bulbo oculare. Il numero varia da una, circolare (pesci e camaleonti), a due (Amnioti), mobili o saldate insieme e trasparenti (alcuni Rettili), a tre. La terza palpebra, o membrana nittitante, è distesa orizzontalmente dall’angolo interno dell’o. o dal disotto della palpebra inferiore.
Dagli Anfibi in poi, il globo oculare possiede varie ghiandole annesse; negli Anfibi si trovano sulla superficie interna della palpebra inferiore; nei Sauropsidi la ghiandola di Harder, o della membrana nittitante, è sita presso l’angolo interno dell’occhio e le ghiandole lacrimali sull’angolo esterno; queste mancano in Coccodrilli, Cetacei e altri Mammiferi acquatici. I margini palpebrali, nei Mammiferi, sono provvisti di ciglia alla cui base sboccano speciali ghiandole sebacee, le ghiandole tarsali o di Meibomio. O. parietale Vescicola di derivazione telencefalica posta sul tetto del diencefalo dei Vertebrati, detta anche organo parietale; differentemente sviluppato nelle diverse classi, è evidente soprattutto negli Squamati e nei Rincocefali, come Sphenodon, dove costituisce un piccolo terzo o. sito nel forame interparietale della volta cranica. Questa vescicola è differenziata distalmente in una porzione lentoide; nella porzione prossimale possiede cellule recettrici a bastoncello (retina eversa) che la collegano al diencefalo. La funzione dell’o. parietale è verosimilmente legata alla capacità di percepire le differenze fra luce e oscurità. In molti Rettili la struttura è sensibile anche all’infrarosso, permettendo così la localizzazione di animali a sangue caldo. O. pineale Altro nome dell’epifisi negli Agnati viventi e in alcuni Elasmobranchi e Squamati, nei quali appare come una vescicola oculiforme. La denominazione si applica propriamente solo nei casi nei quali l’epifisi risulta realmente differenziata in qualche struttura ottica. In tutti gli altri Vertebrati l’organo pineale ha struttura ghiandolare (➔ pineale, ghiandola).
Anatomia umana
Nell’Uomo i caratteri strutturali dell’o. sono pressappoco gli stessi che si osservano negli altri Mammiferi. Il bulbo oculare ha consistenza molle elastica con forma sferoidale. Vi si riconoscono due poli: uno anteriore, detto anche vertice, che corrisponde al centro della cornea, e uno posteriore; la linea che li unisce e che corrisponde al diametro antero-posteriore costituisce l’asse anatomico; per asse ottico si intende invece la linea che parte dal vertice, passa per il centro della pupilla e cade sulla retina, nella regione della visione distinta e cioè in corrispondenza della fovea. Per le distinzioni topografiche si fa riferimento ai meridiani, le circonferenze che passano per i due poli, all’equatore, la circonferenza equidistante dai due poli, e ai piani individuati da tali circonferenze.

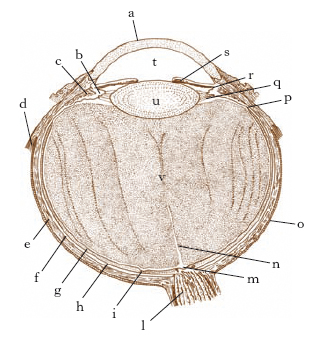
Le pareti del globo oculare sono formate da tre membrane sovrapposte e concentriche (fig. 3): una esterna, fibrosa; una media, a struttura prevalentemente vascolare, contenente anche fibre muscolari lisce e abbondante pigmento; una interna, nervosa. La membrana esterna è costituita da due parti ben distinte: la sclera o sclerotica, membrana connettivale biancastra, opaca e anelastica; e la cornea, anteriore, incastonata nella sclerotica, che costituisce una membrana avvolgente e un mezzo rifrangente. La tunica vascolare, o uvea, consta di tre porzioni, che, procedendo dall’indietro in avanti, sono: la coroide, con funzione nutritizia poiché ricca di vasi sanguigni; il corpo ciliare, formato da elementi vascolari e da fibre muscolari, che costituiscono il cosiddetto muscolo ciliare la cui contrazione determina un aumento della curvatura del cristallino e del suo potere convergente; l’iride, che forma la parte anteriore della tunica vascolare, che traspare attraverso la cornea e si presenta di vario colore nei differenti individui, a seconda della ricchezza in pigmento. L’iride presenta al centro un forame, la pupilla, che, per l’intervento di due ordini di fibre muscolari ad azione antagonista (lo sfintere e il muscolo dilatatore), si restringe sino a circa 2 mm sotto l’azione della luce, dilatandosi invece sino a circa 9 mm con l’oscurità (riflesso fotomotore). La retina o tunica nervosa, tappezza tutta la cavità del bulbo oculare, dal polo posteriore al contorno del foro pupillare. Vi si distinguono due porzioni, che trapassano direttamente l’una nell’altra: il tratto posteriore che costituisce la parte ottica, o retina in senso stretto e il tratto anteriore che prende il nome di parte cieca perché non partecipa alla visione. Carattere comune a tutte le porzioni retiniche è la presenza di un epitelio pigmentato costituito da cellule prismatiche. Sulla superficie interna della porzione ottica della retina vanno segnalati due elementi descrittivi particolarmente importanti: una piccola zona di colorito giallastro in corrispondenza del polo posteriore, la macula lutea (diametro 2-3 mm), recante al centro una depressione, la fovea centralis (diametro ca. 0,3 mm), che è la zona della visione distinta, in cui la sensibilità retinica è massima; una zona rotondeggiante e biancastra, situata medialmente e un po’ inferiormente alla macula: è la papilla ottica, in cui convergono le fibre del nervo ottico e che nella parte centrale presenta una escavazione in cui emergono i vasi destinati alla retina. La papilla costituisce la macchia cieca o punto di Mariotte, in cui mancano i fotorecettori. La retina ha una struttura stratificata a cui partecipano vari elementi morfologici e strutturali: cellule nervose, cellule neurogliche e fibre nervose amieliniche; fra le cellule hanno particolare rilievo, per la loro abbondanza e per la loro funzione fotorecettiva, i coni e i bastoncelli.
Le tre tuniche parietali dell’o. circoscrivono uno spazio interno colmato da mezzi liquidi e solidi, trasparenti e rifrangenti, che i raggi luminosi debbono attraversare prima di poter giungere a stimolare gli elementi sensoriali della retina. Tali mezzi rifrangenti, oltre che dalla cornea, sono rappresentati dall’umor acqueo (con funzione nutritizia e ruolo nella determinazione del tono endooculare), dal cristallino (lente biconvessa al di dietro della pupilla con indice di rifrazione variabile) e dal corpo vitreo (composto da una massa trasparente e gelatinosa, l’umor vitreo, avvolto da un’esile pellicola, la membrana ialoidea).
All’o. sono annesse alcune formazioni anatomiche: la capsula di Tenone, i cosiddetti muscoli estrinseci, la congiuntiva, l’apparato lacrimale, le palpebre, le sopracciglia.
Biochimica
Le varie parti che costituiscono l’o. sono caratterizzate da un’elevata specificità biochimica e metabolica. La sclera e la congiuntiva sono costituite da fibre di collageno e da mucopolisaccaridi. La sclera contiene condroitinsolfato A, B e C ed è opaca perché le sue fibre di collageno sono variamente intrecciate. La cornea, viceversa, è costituita da fibre di collageno che decorrono parallelamente fra loro nei diversi strati e ne causano la trasparenza. Lo stroma della cornea è costituito da tessuto connettivo di origine mesenchimale il cui principale glicosamminoglicano è il cheratansolfato. La superficie esterna della cornea è rivestita da un epitelio polistratificato, impermeabile alle lacrime, riccamente innervato, che possiede un’elevata concentrazione di acetilcolina e degli enzimi deputati al suo metabolismo (acetilcolinesterasi e colina acetiltransferasi). Una delle caratteristiche principali del metabolismo corneale è la preponderanza della via dei pentosofosfati attraverso cui viene degradato circa il 50% del glucosio utilizzato dal tessuto. Le elevate concentrazioni di nucleotidi piridinici e l’elevato consumo di O2 indicano un’intensa attività metabolica delle cellule corneali.
La fonte principale di nutrimento è rappresentata dall’umore acqueo, nel quale sono presenti i substrati e l’ossigeno per il metabolismo. Il cristallino è costituito da un singolo strato epiteliale di origine ectodermica e dalle fibre del cristallino. La capsula del cristallino contiene un collageno ricco di idrossilisina, i cui gruppi δ-idrossilici sono uniti a glucosilgalattosio mediante legami O-glicosidici. Tra le principali proteine del cristallino l’α-cristallina costituisce circa il 75% delle proteine totali. Si tratta di una proteina formata da due catene polipeptidiche di 173 (A) e 175 (B) amminoacidi che durante l’invecchiamento sono modificate per proteolisi con perdita dei residui terminali, formando aggregati proteici che causano un aumento dell’opacità del nucleo centrale del cristallino. Il metabolismo del cristallino è relativamente basso ed è basato principalmente sulla glicolisi e sulla via dei pentosofosfati. Una caratteristica del cristallino è quella di possedere concentrazioni molto elevate dei composti antiossidanti glutatione (600 mg/100 g di corteccia) e acido ascorbico (30 mg/100 g di tessuto). Inoltre, sono presenti analoghi del glutatione (acido oftalmico, acido noroftalmico, S-solfoglutatione) nei quali la cisteina è sostituita da un diverso amminoacido.
Nella cateratta senile, oltre alla già citata alterazione delle proteine si osserva un iniziale aumento dell’idratazione e successiva riduzione del liquido. Oltre all’aumento di Na+ e alla riduzione di K+ si verifica anche un aumento di Ca++, parametro che è stato utilizzato per classificare gli stadi di sviluppo della cataratta. Nella cateratta diabetica si osserva un accumulo di fruttosio e di sorbitolo nel cristallino; il metabolismo del glucosio viene inibito e la concentrazione di ATP e la sintesi proteica si riducono.
La camera interna dell’o. è rivestita dalla melanina dell’epitelio pigmentato e della struttura coroidea, posta tra il tessuto nervoso della retina e la sclera. La melanina è la sostanza che conferisce colore anche all’iride. La retina è il tessuto in cui si verifica il maggior consumo di ossigeno per unità di massa. Il metabolismo cellulare è prevalentemente aerobico, anche se la glicolisi è fondamentale per l’espletamento della funzione visiva. Nella retina si verificano le reazioni fotochimiche della visione alle quali partecipano tutti i pigmenti visivi.
Per il meccanismo biochimico della visione ➔ rodopsina.
Fisiologia
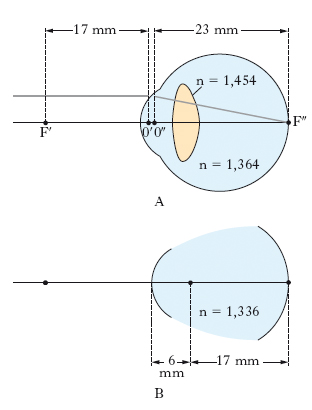
L’o. è sede dei processi iniziali della visione, grazie al suo sistema diottrico che forma sulla superficie retinica l’immagine luminosa degli oggetti dell’ambiente esterno; con la intermediazione delle conseguenti reazioni fotochimiche che si svolgono a livello dei recettori retinici, l’o. trasforma gli impulsi luminosi in stimoli elettrici e li inoltra alle vie ottiche. Come sistema ottico, l’o. normale o emmetropico è un sistema diottrico e corretto il cui secondo fuoco cade sulla retina, atto cioè a formare esattamente, su quest’ultima, l’immagine stigmatica, ortoscopica, acromatica di un oggetto luminoso posto all’infinito. In condizioni anormali (ametropia), per alterato potere convergente dei mezzi rifrangenti o per abnormi dimensioni del diametro antero-posteriore dell’o., i raggi luminosi paralleli provenienti da un oggetto all’infinito possono formare il loro fuoco al di là della retina (ipermetropia), o davanti a essa (miopia). Nell’o. normale, l’angolo di campo corrispondente alla visione distinta, cioè corrispondente al formarsi dell’immagine sulla fovea, è solo di circa 1°, ma la mobilità del bulbo oculare nell’orbita consente la visione distinta entro un angolo di circa 70°; l’angolo di campo complessivo della visione indistinta è di circa 160°. Nella fig. 4 sono riportate le posizioni dei fuochi F′, F″ e dei punti principali O′, O″ del sistema ottico centrato equivalente all’o. normale (o. schematico: fig. 4A), nonché le dimensioni del diottro sferico equivalente (o. ridotto o o. di Listing: fig. 4B). Dei vari mezzi diottrici, la cornea, l’umore acqueo e il vitreo sono inerti; il cristallino possiede invece la capacità di far variare il suo potere convergente e di svolgere una funzione accomodativa, che garantisce la precisa messa a fuoco dell’immagine sulla retina, quale che sia la distanza dell’oggetto.
Quando i raggi luminosi colpiscono la retina determinano nel pigmento contenuto nei fotorecettori (i coni – preposti alla visione diurna, alla sensibilità cromatica, alla discriminazione spaziale – e i bastoncelli che assicurano la visione crepuscolare e registrano le variazioni di intensità della luce) dei profondi cambiamenti biochimici che concernono sia il gruppo cromoforo sia la proteina che gli è associata. Contemporaneamente a questi mutamenti si destano nel recettore dei potenziali che si propagano lungo la fibra del nervo ottico che è emanazione della catena.
Patologia
Le anomalie di sviluppo dell’o. possono interessare l’organo in toto (anoftalmia, microftalmia ecc.) o parti di esso (aniridia, acoria, coloboma ecc.), o i suoi annessi (soprattutto le palpebre: blefarofimosi, microblefaria ecc.).
L’apparato oculare può essere colpito, oltre che da molteplici lesioni traumatiche (da cause fisiche, chimiche, radianti), da vari processi morbosi. Essi possono interessare l’o. umano nel suo insieme (panoftalmite, glaucoma) o suoi elementi strutturali: la cornea (cheratiti), l’uvea (uveiti), la retina (retinopatie, distacco retinico), il cristallino (cataratta) ecc. Relativamente frequenti sono i disturbi della mobilità oculare (strabismo), i vizi di rifrazione dell’o. (miopia, ipermetropia, astigmatismo), i processi infiammatori a carico degli annessi oculari: congiuntiva (congiuntivite), palpebre (blefariti), lacrimali (dacriocistite). La patologia oculare è di pertinenza strettamente oculistica.
Botanica e agraria
In certi frutti con ovario infero (pera, mela), la nicchia che si trova all’apice del frutto e che corrisponde all’apertura della coppa ricettacolare, la quale è concresciuta con i carpelli. O. è anche sinonimo di gemma dormiente.
È detta o. di pavone (o vaiolo dell’olivo) una malattia dell’olivo, dovuta al fungo Deuteromicete Cycloconium oleaginum: sulle foglie compaiono macchie rotonde, che presentano zone concentriche alternatamente chiare e scure, le quali ricordano gli ocelli delle penne del pavone. La malattia, diffusa ma poco dannosa, determina la caduta delle foglie.
Fisica
O. fotometrico normale
Cellula fotoelettrica con curva spettrale di visibilità relativa coincidente con quella rappresentante mediamente, per convenzione internazionale, un o. umano normale a cui ci si riferisce in alcune misurazioni fotometriche.
In ottica, obiettivo a o. di pesce (fish-eye), obiettivo grandangolare con angolo del campo molto grande, tra 140° e 180°.
Meteorologia
O. di un ciclone
Zona di calma al centro del ciclone, dove la pressione atmosferica raggiunge il minimo valore.
Religione
Il simbolismo religioso dell’o., che si trova diffuso in forme varie e presso molti popoli antichi e moderni, deriva soprattutto da due esperienze umane distinte: il valore conoscitivo del vedere e la potenza fascinatrice dello sguardo. La divinità onnisciente è concepita dovunque come onniveggente e spesso rappresentata come dotata di numerosi o.; l’immagine di un solo o., eventualmente incluso in un triangolo (trinità), ricorda l’onniveggenza di Dio. Nelle religioni che conoscono un dio celeste, spesso il sole, la luna, le stelle sono o. di Dio. Gli esseri demoniaci, fra altri tratti mostruosi, hanno spesso qualche anormalità dell’o.: hanno un solo o. (per es., i Ciclopi), vedono più di notte che di giorno ecc. Alla potenza dello sguardo si legano le credenze nel malocchio contro il quale molti popoli ricorrono ad amuleti in forma di occhio.
Tecnica
O. di biella
Cerniera del piede di biella, in cui è alloggiato lo spinotto.
O. magico
Indicatore ottico di sintonia per radioricevitori, costituito da un tubo termoelettronico (a sua volta composto di un triodo e di un minuscolo e semplificato tubo a raggi catodici).